PROKARIOTIK
Gbr. Penampang Sel Prokariotik
Sel prokariotik adalah sel yang belum mempunyai inti sejati. Ciri-ciri sel prokariotik adalah bahan genetik (DNA) tidak terstruktur dalam bentuk nucleus, DNA terdapat pada nukleoid yang tidak diselubungi oleh membran.
Secara umum sel prokariotik memiliki ukuran yang lebih kecil dibandingkan sel eukariotik. Setiap prokariotik merupakan sel tunggal, tetapi akan sering terlihat dalam tipe rantai, agregat, atau kelompok sel yang jumlahnya ratusan.
SEL EUKARIOTIK
Gambar : penampang sel eukariotik
Sel eukariotik adalah sel yang mempunyai inti sel sejati. Sel eukariotik tersusun atas:
· Membran sel
Struktur terluar sel berupa membran sel. Membrane tersusun atas lapisan lipoprotein (gabungan antara lemak dan protein pada membran sel darah merah, 50%lemak-50% protein).
· Sitoplasma
Cairan yang terdapat dalam sel dan terletak di luar inti sel. Cairan yang terdapat di inti sel disebut nukleoplasma. Matrik sitoplasma terdapat dalam lingkungan dalam sel. Matrik sitoplasma berfungsi mengatur kekentalan sitoplasma, pembentukan benang spindel, pembentukan berbagai serabut (mikrotubulus, filamen, myofibril).
· Organel sel
1. Mitokondria (terdapat di semua sel eukariotik)- berfungsi sebagai pembuat dan pemberi energy
2. Peroksisom (badan mikro)Peroksisom dan glikosom berasosiasi membentuk badan mikro (struktur serupa dengan lisosom). Terletak di dekat mitokondria atau kloroplas. Berfungsi menghasilkan enzim katalase (berfungsi menguraikn peroksida hydrogen)
3. Mikrotubulus (berfungsi untuk membentuk silia, sentriol dan benang-benang spindel)
4. Mikrofilamen (tersusun atas untaian protein globular,protein kontraktil. Bersama myosin menghasilkan kontraksi)
5. Nucleus (inti sel, tempat berlangsung sintesis molekul RNA)
6. Retikulum Endoplasma RE kasar(permukaan melekat pada ribosom)Re halus(tidak terdapat ribosom.Fungsi: detoksifikasi, mengangkut golgi komplek, melaksanakan oksidasi awal lemak, menyusun fosfolipid, gilkolipid, dll)
7. Aparatus Golgi (kumpulan sisternae, yang terletak di sel tumbuhan-diktiosom. Dapat membentuk lisosom)
8. Ribosom (fungsi utama, mensintesis protein)Ribosom yang menempel 1 dengan yang lain disebut polisom, diikat oleh mRNA.
9. Lisosom (berfungsi untuk pencernaan, menghasilkan antibody)Lisosom primer,memproduksi sel yang belum aktif. Berfungsi sebagai vakuola makanan)Lisosom sekunder,terlibat dalam kegiatan mencerna. Berfungsi sebagai autofagosom.
10. Kloroplas (plastisida yang mengandung pigmen hijau-klorofil. Tempat berlangsungnya fotosintesis)
11. Sentrosom
12. Dinding sel (tersusun atas selulosa dan devirat-deviratnya. Berfungsi untuk menjaga bentuk sel agar tetap dan sebagai proteksi sel terhadap gangguan mekanis)
13. Vakuola Vakuola kontraktilVakuola non-kontraktil
Struktur Sel Prokariotik dan Eukariotik
Sejak ditemukannya mikroskop elektron para ahli biologi mulai berhasil mengidentifikasi struktur internal dari berbagai macam sel. Berdasarkan hasil pengamatannya, para ahli menggolongkan sel menjadi dua kelompok, yaitu sel prokariotik dan sel eukariotik. Penggolongan ini didasarkan atas ukuran dan struktur intemal atau kandungan organel selnya. Sel prokariotik memiliki struktur yang sederhana,. misalnya bakteri, ganggang hijau-biru, dan mikoplasma. Sedangkan, sel eukariotik memiliki struktur yang lebih kompleks, misalnya protista, fungi, tumbuhan, dan hewan.
1. Struktur Sel Prokariotik
Prokariotik meliputi archaebakteria (bakteri purba) dan eubakteria (bakteri modern/bakteri sejati) yang beranggotakan bakteri, mikoplasma dan alga hijau-biru. Ukuran sel prokariotik berkisar antara 0,5 -3 mm. Struktur umum sel prokariotik yang diwakili oleh bakteri berturut-turut mulai dari luar ke dalam adalah dinding sel, membran sel, mesosom, sitoplasma, ribosom dan materi inti (DNA dan RNA).
Dinding sel bakteri berfungsi untuk menahan tekanan osmotic sitoplasma, sehingga sel tidak mudah pecah akibat masuknya air kedalam sel, dinding sel bakteri tersusun atas peptidoglikan atau mukopepetida yang dapat dipergunakan sebagai dasar penggolongan bakteri menjadi dua golongan , yaitu bakteri gram positif dan bakteri gram negative. Pada bajteri gram positif, hamper 90% komponen dinding selnya tersusun atas peptidoglikan, sedangkan pada bakteri gram negative berkisar antara 5 – 20%.
Selaput sitoplasma atau membran sel bakteri berfungsi dalam seleksi dan pengangkutan larutan ke dalam sel; berperan dalam transfer elektron dan fosforilasi oksidatil; pada bakteri aerob berperan dalam pengeluaran enzim hidrolitik; sebagai tempat enzim dan molekul pembawa yang berfungsi dalam biosintesis DNA, polimer dinding sel dan lipid selaput
Komponen utama membran sel tersusun atas lipid dan protein atau lipoprotein. Membran sel bakteri dan sianobakteri membentuk lipatan ke dalam yang dinamakan mesosom. Pada beberapa bakteri, mesosom berperan dalam pembelahan sel. Sedangkan pada sianobakteri, mesosom berfungsi sebagai kompleks fotosintetik yang mengadung pigmen fotosintesis.
Di dalam sitoplasma terdapat kurang lebih 20.000 - 30.000 ribosom yang tersusun atas RNA dan protein. Ribosom merupakan tempat sintesis protein. Ribosom prokariotik tersusun atas sub unit kecil dan sub unit besar yang berukuran 30 S dan 50 S (Svedberg). Pada saat proses transaksi, kedua sub unit ini bersatu untuk menjalankan fungsinya. Di dalam sitoplasma juga terdapat molekul protein dan enzim yang digunakan dalam setiap reaksi kimia di dalam sitoplasma. Bakteri juga menyimpan cadangan makanan di sitoplasma dalam bentuk granula-granula tidak larut air. Materi genetik sel prokariotik membentuk suatu struktur yang dinamakan nukleoid, merupakan kromosom tunggal. Antara materi inti dengan sitoplasma tidak terdapat pembatas atau tidak memiliki membrane inti. Sel prokariotik mengandung sejumlah kecil DNA dengan total panjang antara 0,25 mm sampai 3 mm yang mampu mengkode 2000 – 3000 protein.
2. Struktur Sel Eukariotik
Sel eukariotik biasanya merupakan penyusun struktur makhluk hidup multi seluler. Sel eukariotik tersusun atas membrane sel, sitoplasma, nukleus, sentriol, retikulum endoplasma, ribosom, komplek golgi, lisosom, badan mikro, mitrokondria, mikrotubulus dan mikro filamen. Organelorganel di dalam sel memiliki peran yang sangat penting bagi kelangsungan hidup sel tersebut. Setiap organel di dalam sel memiliki fungsi yang berbeda - beda.
1. Membran Sel
Sel memiliki struktur khusus yang berfungsi untuk memisahkan isi sel dengan lingkungan luarnya, struktur ini dinamakan membrane plasma atau membran sel. Membran plasma ini memiliki ketebalan antara 5 sampai 10 nm (nanometer), oleh karena itu hanya dapat dilihat dengan mikroskop elektron
. Membran sel memiliki beberapa fungsi, antara lain yaitu:
1) Sebagai pembungkus isi sel dan membentuk sistem endomembran di dalam sel, misalnya retikulum endoplasma, aparatus Golgi, dan lisosom.
2) Menyediakan selaput atau penghalang yang bersifat selektif permeabel. Membran sel berfungsi untuk menyaring masuknya zat-zat ke dalam sel sehingga tidak semua zat dapat menembus membran sel.
3) Sebagai sarana transpor larutan dari dan ke dalam sel. Membran sel berfungsi dalam membantu memasukkan dan mengeluarkan senyawa – senyawa tertentu dari dan ke dalam sel.
4) Merespons terhadap sinyal dari luar. Pada membran sel terdapat protein integral yang berfungsi sebagai reseptor untuk menerima sinyal dari lingkungan sel.
5) Untuk interaksi interseluler. Protein - protein membran sel dan glikoprotein sebagai perantara sel untuk berinteraksi dengan sel lain atau dengan lingkungan luarnya.
6) Tempat aktivitas biokimiawi. Beberapa reaksi kimia dikatalisis oleh protein integral membran yang berfungsi sebagai katalisator.
7) Untuk transduksi energi. Membran dalam (inner membrane) kloroplas berfungsi untuk mengubah energi cahaya menjadi energi kimia dalam proses fotosintesis.
Semua membran sel terdiri atas dua komponen utama, yaitu lemak (lipid) dan protein yang terikat secara non kovalen dan tersusun dalam suatu struktur yang menyerupai lembaran. Lembaran tersebut tersusun atas dua lapisan lemak yang dinamakan lipid bilayer. sedangkan protein terletak di antara lemak atau di permukaan lapisan lipid bilayer.
Perbandingan jumlah, antara lemak dan protein bervariasi, tergantung dari jenis membran sel, misalnya membran retikulum endoplasma berbeda dengan membran Golgi, jenis organisme, misalnya membran sel tumbuhan berbeda dengan membran sel hewan dan jenis sel, misalnya membran sel tulang berbeda dengan sel hati. Karbohidrat terikat secara kovalen, baik dengan lemak maupun protein.
Karbohidrat yang terikat dengan lemak dinamakan glikolipid, sedangkan yang terikat dengan protein dinamakan glikoprotein. Baik glikolipid maupun glikoprotein berfungsi sebagai media interaksi dengan sel lainnya. sekitar 2 - 10 ppersen glikolrotein membangun membran plasma, tergantung dari tipe sel dun spesies. Fungsi glikolipid masih belum banyak diketahui, tetapi diduga berhubungan dengan tempat melekatnya beberapa mikroorganisme infektif. Kolesterol pada membran plasma hanya dijumpai pada sel hewan dan sekitar 50% dari lemak membran terdiri atas kolesterol. Fungsi kolesterol pada membrane berhubungan dengan rigiditas atau kekakuan membran.
Protein yang menyusun membran plasma tersusun atas lebih dari 50 jenis protein yang berbeda. |enis-jenis tersebut terletak dengan orientasi tertentu pada lipid bilayer. Protein membran dikelompokkan menjadi tiga jenis, yaitu:
1) Protein integral
Protein ini menembus lipid bilayer sehingga memiliki dua permukaan, yaitu permukaan yang mengarah ke lingkungan luar sel dan yang menghadap ke dalam sitoplasma
2) Protein perifer
Protein ini terdapat pada permukaan luar lipid bilayer atau pada permukaan dalam-lipid bilayer. Ikatan antara protein perifer dengan lipid bilayer adalah non kovalen.
3) Protein yang terikat lipid membran
Protein ini terikat secara kovalen dengan lipid bilayer dan terletak pada permukaan luar dari lipid bilayer.
b. Sitoplasma
Sitoplasma merupakan cairan sel yang dibungkus oleh membrane plasma. sitoplasma mengandung gula, asam amino, lemak,ion-ion dan senyawa kimia lain yang digunakan untuk metabolisme sel. Di dalam sitoplasma terdapat membran intrasel yang membungkus organel sel, misalnya membran yang membungkus mitokrondria, kloroplas, lisosom, peroksisom, retikulum endoplasma, dan badan Golgi.
Bagian sitoplasma yang berada di antara organel dinamakan sitosol. Volume sitosol lebih kurang 50% dari volume sel. Di dalam sitosol juga terdapat protein dan enzim-enzim untuk reaksi kimia.
c. Mitokondria
Ukuran mitokondria bervariasi, tetapi rata-rata ukuran diameternya antara 0,2 - 0,7 mikrometer (pm) dan panjangnya antara 1 - 4 mikrometer.
Ukuran mitokondria ini hampir sama dengan ukuran bakteri yang menunjukkan salah satu bukti evolusi bahwa mitokondria merupakan bakteri yang bersimbiosis dengan sel eukoriotik. Bentuk mitokondria bervariasi, tergantung dari jenis selnya, misalnya pada sel-sel awal embrio, bentuk mitokondrianya bulat atau oval, sedangkan pada sel-sel lain bentuknya seperti gelendong dan ada juga yang berbentuk pipa. Karena ukurannya yang relatif besat mitokondria dapat terlihat cukup jelas di bawah mikroskop cahaya. Pada umumnya, mitokondria tersebar secara acak di dalam sel dan cenderung berkumpul pada bagian sel yang banyak memerlukan energi, misalnya di sekitar gelendong pembelahan, atau di sekitar memmbran yang melakukan endositosis. Jumlah mitokondria di dalam sel bervariasi tergantung dari jenis sel, spesies organisme, dan keadaan fisiologi sel. Selsel yang metabolismenya aktif banyak mengandung mitokondria dibandingkan sel-sel yang tidak aktif.
Mitokondria memiliki kelenturan yang tinggi sehingga bentuknya dapat berubah-ubah dari waktu ke waktu. Selain itu, mitokondria mampu bergerak atau berpindah dari satu tempat ke tempat lain dalam sitoplasma.
Bagian-bagian utama mitokondria dibedakan menjadi dua, yaitu bagian selaput atau membran dan bagian matriks. Membran mitokondria ada dua yaitu membran luar dan membran dalam. Antara membran dalam dan membran luar terdapat ruang antarmembran yang berisi berbagai macam enzim. Membran luar mitokondria lebih tipis dari pada membrane dalam yaitu kurang dari 6 nanometer, sedangkan membran dalam berukuran antara 6 - 8 nanometer. Membran dalam mitokondria membentuk juluran-juluran ke arah matrik sehingga memperluas permukaan dalamnva. Iuluran membran ke arah matriks ini dinamakan tristae. Matriks mitokondria merupakan bagian mitokondria yang
menyerupai gel. Di dalam matriks mitokondria terdapat ribosom, DNA, RNA dan beberapa protein yang larut dalam air serta filamen, dan granul.
Pada membran dalam (inner membrane) mitokondria terdapat beberapa jenis protein yang terlibat dalam proses pembentukan ATP. Di dalam sel, ATP merupakan molekul berenergi tinggi yang akan digunakan untuk metobolisme sel. Selain berfungsi menghasilkan energi dalam bentuk ATP, mitokondria juga berfungsi sebagai tempat penyimpanan ion kalsium di dalam sel. Ion-ion ini disimpan dalam suatu badan khusus yang dinamakan
granul. Mitokondria di dalam sel mampu menggandakan diri, sehingga jumlahnya dapat bertambah sesuai dengan kebutuhan energi sel.
d. Retikulum Endoplasma
Gbr. Retikulum Endoplasma (RE Kasar dan RE Halus)
Retikulum Endoplasma (RE) merupakan bentukan membran yang sangat berlipat-lipat membatasi suatu ruangan yang disebut lumen (sisterna). Antara lumen RE dengan sitosol hanya dipisahkan oleh selapis membran sehingga memudahkan terjadinya pertukaran zat antara lumen RE dengan sitosol. Berdasarkan ada tidaknya ribosom yang menempel pada permukaan luar membran, RE dibedakan menjadi dua, yaitu Retikulum Endoplasma Halus (Smooth Endoplasmic Reticulumi /SER) dan Retikulum Endoplasma Kasar(Rough Endoplasmic Reticulum / RER). Pada RER permukaan luar membrannya banyak ditempeli oleh ribosom.sebaliknya pada SER permukaan luar membrannya tidak ditempeli oleh ribosom. RER banyak dijumpai pada sel-sel yang aktif mensekresikan protein misalnya sel – sel pancreas, kelenjar ludah, dan kelenjar lainnya.
Protein yang dihasilkan dari RER antara lain adalah protein yang disekresikan keluar sel, protein integral membran, protein-protein khusus di dalam organel, seperti protein di dalam Golgi, lisosom, endosom, dan vakuola, makanan pada sel tumbuhan. SER banyak ditemukan pada otot rangka, tubulus ginjal, dan kelenjar endokrin yang mensekresikan hormon steroid.
SER mempunyai beberapa fungsi, yaitu:
• Sintesis hormon steroid pada sel-sel kelenjar endokrin pada gonad dan adrenal.
• Detoksifikasi di dalam hati yang melibatkan beberapa molekul penting di dalam sel hati.
• Melepaskan glukosa dari glukosa-6-fosfat di dalam sel-sel hati.
• Sebagai tempat melekatnya granul-granul yang berisi glikogen pada sel-sel hati.
• Tempat menyimpan ion-ion kalsium di dalam sisterna yang akan dikeluarkan jika ada rangsangan yang menyebabkan pengeluaran ion kalsium, misalnva kontraksi otot.
e. Aparatus Golgi atau Kompleks Golgi.
Aparatus Golgi (AG) atau Kompleks Golgi pertama kali ditemukan oleh Camilio Golgi tahun 1898 di dalam sitoplasma sel saraf. AG dijumpai hampir pada semua sel tumbuhan dan sel hewan.
Organel ini terdiri atas setumpuk saku-saku pipih yang masing-masing dibatasi oleh selapis membian. Dengan menggunakan mikroskop elektron, tampak bahwa AG tersusun atas tiga bentukan membran, yaitu:
1) kantung-kantung pipih yang disebut sisterna atau sakulus, kantung – kantung pipih tersebut tersusun bertumpuk membentuk diktiosom,
2) vesikel-vesikel kecil berdiameter kurang lebih 50 mikrometer yang terletak pada sisi yang berbatasan dengan RE, vesikel ini dinamakan vesikel tiansisi atau vesikel peralihan, fungsi vesikel adalah membawa protein dan lipid dari RE ke AG dan dari sakulus satu ke sakulus lainnya,
3) vesikel besar yang terletak pada sisi yang berhadapan dengan membrane plasma, vesikel ini dinamakan vesikel sekretori,vesikel sekretori adalah membawa protein atau lipid yang telah mengalami Pemrosesan di dalam lumen sakulus.
Beberapa penelitian membuktikan bahwa AG tidak hanya berfungsi sebagai alai transport materi ke luar sel. Akan tetapi banyak reaksi yang berlangsung di dalam lumen AG, antara lain proses biosintesis- glikoprotein dan glikolipid yang dikatalisis oleh enzim glikosil transferase, kedua proses ini sering dinamakan glikosilasi. Di dalam AG juga terjadi proses penambaKan gugus sulfat pada karbohidrat yang dikatalisis oleh enzim sulfat tansferase. Seiain itu, di dalam lumen AG terjadi proses sintesis proteoglikan yang merupakan komponen matriks ekstra sel. Pada sel tumbuhan yang sedang membelah, AG berperanan dalam pembentukan komponen dinding sel yang baru.
Molekul-molekul protein dan lipid yang telah mengalami modifikasi kimiawi di dalam lumen AG akan di packing oleh membran Golgi dan ditransfer dalam bentuk vesikel. ‘
Ada tiga macam protein yang dihasilkan oleh Golgi, antara lain:
1) protein membran inti, membran plasma dan protein membran organel
2) protein sekretori yang disimpan dalam bentuk vesikel
3) protein enzim yang disimpan dalam vesikel (lisosom)
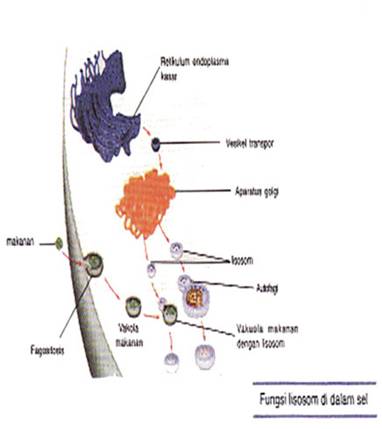
f. lisosom
Lisosom pertama kali ditemukan pada tahun 1949 oleh De Duve di dalam serpihan sel-sel hati. Organel ini berbentuk semacam kantung yang berisi enzim hidrolitik. Selama masih terbungkus membran, enzim hidrolitik bersifat stabil. Terdapat lebih kurang 40 macam enzim hidrolitik yang ditemukan di dalam lisosom. Enzim-enzim tersebut meliputi protease,nuklease, glikosidase, lipase, fosfolipase, fosfatase dan sulfatase. Enzim – enzim tersebut hanya akan dapat bekerja optimal pada pH sekitar 5.membran lisosom mengandung protein transfer untuk membawa hasil pencernaan ke sitosol. Membran lisosom tidak akan tercerna oleh enzim yang dikandungnya sendiri karena kandungan karbohidrat yang tinggi pada membrannva.
Lisosom tergolong organel yang polimorfik karena memiliki bentuk dan ukuran yang bervariasi. Ada empat macam bentuk lisosom, yaitu satu macam lisosom primer dan tiga macam lisosom sekunder. Lisosom primer adalah lisosom yang baru terbentuk dari AG dan belum berfusi (bergabung) dengan materi yang akan dicerna. Lisosom sekunder ada tiga macam,yaiitu:
(1) heterofagosom, merupakan gabungan antara lisosom primer dengan fagosom,
(2) Sitolisosom merupakan gabungan antara lisosom primer dengan autosom,
( 3) badan residu, adalah vakuola yang berisi sisa materi yang tidak tercerna
Fungsi utama lisosom adalah untuk pencernaan intra sel. Materi yang dicerna oleh lisosom dapat berasal dari luar sel atau dari dalam sel itu sendiri. Materi dari luar sel masuk ke dalam sitoplasma melalui pinositosis dan fagositosis. Pencernaan intra sel selalu terjadi di dalam lisosom, enzim, hidorolitik tidak pernah keluar dari dalam lisosom sehinggan pencernaan berlangsung optimal. Akan tetapi, jika membran lisosom pecah, maka enzim hidrolitik pada lisosom akan keluar dan mencerna sel itu sendiri.
Beberapa peran lisosom antara lain adalah:
1) perombakan organel sel yang telah tua
2) proses metamoifosis pada katak, misalnya menyusutnya ekor pada berudu karena dicerna oleh enzim katepsin di dalam lisosom
3) pemulihan ukuran uterus setelah kehamilan
4) proses fertiliasi, dimana bagian kepala sperma yang dinamakan akrosom mengandung enzim hialuronidase untuk mencerna zona pelusida pada sel telur.
Hasil pencernaan lisosom, seperti asam amino, glukosa dan nukleotida mampu menembus membran lisosom menuju sitosol. Membran lisosom selanjutnya akan dikembalikan menuju membran plasma melalui proses eksositosis. pencernaan bagian - bagian sel yang telah tua dinamakan autofagi.
g. Badan Mikro
1) Peroksisom
Organel ini ditemukan pada sel hewan, sel tumbuhan tertentu maupun sel ragi. Peroksisom pertama kali ditemukan oleh De Duve dan kawan-kawannya pada tahun 1965 di dalam sel-sel hati. Di dalam peroksisom ditemukan beberapa macam enzim oksidase dan enzim katalase. Oleh karena enzim - enzim ini berperan dalam pembentukan katalase. oleh karena enzim - enzim ini berperan dalam pembentukan dan pembongkaran hidrogen peroksida(H2O2) , maka organel tersebut dinamakan peroksisom.Pada sel tumbuhan, fungsi organel ini berkaitan dengan siklus glioksilat sehingga dinamakan glioksisom.
Di dalam sel, peroksisom berbentuk bulat telur dengan diameter kurang lebih antara 0,5 - 0,7 mikrometer, hanya dibungkus oleh selapis membran. Jumlah peroksisom untuk tiap sel bervariasi antara 70-700. Peroksisom memiliki kemampuan untuk membelah diri sehingga dapat membentuk peroksisom anak. Protein dan lipid yang diperlukan
ditransfer dari sitosol. Selain berfungsi untuk pembentukan dan perombakan H2O, menjadi substrat organik dan H2O, peroksisom juga berfungsi untuk merombak asam lemak yang tersimpan dalam biji menjadi glukosa untuk proses perkecambahan.
2) Glioksisom
Glioksisom merupakan badan mikro yang hanya ditemukan pada sel tumbuhan. Diameter glioksisom antara 0,5 sampai 1,0 mikrometer. Sedangkan peroksisom merupakan badan mikro yang ditemukan baik pada sel hewan maupun sel tumbuhan. Glioksisom banyak ditemukan pada biji-bijian yang berperan sebagai tempat menyimpan asam lemak untuk pembentukan energi dalam Proses perkecambahan.
Salah satu proses utama pada biji yang sedang mengalami perkecambahan adalah perubahan dari asam lemak dalam glioksisom, menjadi karbohidrat atau disebut glukoneogenesis. Penguraian asam lemak menjadi asetil ko-A selanjutnya berubah menjadi oksaloasetat untuk membentuk sitrat. Asam sitrat yang terbentuk akan diubah menjadi glukosa melalui serangkaian reaksi enzimatis yang terdapat di dalam glioksisom.
h.Ribosom
Ribosom merupakan salah satu organel tidak bermembran yang ditemukan pada semua sel, baik sel prokariotik maupun eukariotik. Pada eukariotik , organel ini terdapat pada sitoplasma, menempel pada permukaan luar retikulum endoplasma, didalam metriks mitokondria dan didalam stroma kloroplas.
Ribosom terdiri atas dua sub unit yaitu sub unit besar darn sub unit kecil. Kedua sub unit ini akan berfusi jika proses trnaslasi berlangsung.Sub unit ribosom dinyatakan dengan satuan S (Svedberg) yang merupakan nama penemunya, satuan ini menunjukkan kecepatan pengendapan pada saat sub unit tersebut disentrifugasi, misalnya sub unit kecil dan sub unit.
besar ribosom pada eukariotik adalah 40S dan 60s. Komponen penyusun besar ribosom terdiri atas protein ribosom dan ARN ribosom (ARN-r). Protein ribosom disintesis oleh bebas yang terdapat di dalam sitoplasma, sedangkan ARN-r ditranskripsi di dalam anak inti (nukleous).
Organel ini merupakan tempat berlangsungnya penerjemahan (translasi) kodon (kode genetik) yang dibawa ARN-duta (ARN-d). Hasil translasi ini adalah polipeptida. Polipeptida hasil translasi pada RER akan dikirim dan diolah di dalam AG menjadi protein membran, dan enzim lisosom, atau disekresikan ke luar sel melalui vesikel. Sedangkan polipeptida
hasil translasi pada ribosom bebas dikirim ke mitokondria, sebagai enzim peroksisom, atau sebagai protein ribosom.
i. Sitoskeleton
Di dalam sitosol juga ditemukan adanya sitoskeleton yang tersusun atas mikrotubulus, mikrofilamen dan filamen intermediat. Sitoskeleton berfungsi untuk menyokong bentuk sel dan memungkin terjadinya gerakan-gerakan organel di dalam sitoplasma. Mikrotubulus ada yang Ietaknya terbenam di dalam sitosol, dinamakan mikrotubulus sitoplasmik
dan ada juga yang berfungsi sebagai penyusun organel , sepe’rti silia, flagela,
dan sentriol. Mikrofilamen merupakan protein konEaktil yang berfr-rngsi
untuk pergerakan di dalam sitoplasma, misalnya aliran sitoplasma cii clalanr
sel tumbuhan dan gerak amoeboid pada leukosit.
1. Mikrotubulus
Mikrotubulus tersusun atas molekul protein tubulin. Ada dua jenis protein tubulin penyusun tubulin, yaitu tubulin α dan tubulin β. Setiap mikrotubulus tersusun atas 13 protofilamen yang tersusun paralel mengelilingi suatu sumbu. Ada dua macam mikrotubulus di dalam sel yang dibedakan atas stabilitasnya, yaitu mikrotubulus stabil dan mikrotubulus labil. Contoh mikrotulus stabil adalah pembentuk silia dan flagela. Sedangkan mikrotubulus labil contohnva mikrotubulus pembentuk gelendong pembelahan.
Mikrotubulus sitoplasmik didalam sel berfungsi sebagi keranga dalam yang menetukan bentuk sel dan untuk transfer molekul di dalam sel. Mikrotubulus ini berbentuk serabut tunggal dengan diameter lebih kurang 25 nanometer. Beberapa organel yang tersusun dari mikrotubulus adalah sentriol, silia dan flagella.
2. Mikrofilamen
Mikrofilamen biasanya banyak terdistribusi dibawah permukaan membrane plasma. Panjang mikrofilamen bervariasi, dengan diameter lebih kurang 7 µm. Mikrofilamen tersusun atas protein, terutama aktin dan miosin. Hampir semua jenis sel hewan mengandung aktin. Aktin dan miosin banyak ditemukan terutama pada sel otot, dengan komposisi miosin yang lebih sedikit dibandingkan aktin. Kedua jenis protein ini berperan untuk pergerakan, misalnya aliran sitoplasma pada sel tumbuhan (siklosis), dan gerak amoeboid pada Protozoa.
3. Filamen lntermediet
Filamen intermediet memiliki diameter antara 8-10 pm, berbentuk pembuluh, tersusun atas 4-5 protofilamen yang tersusun melingkar, bersifat liat, stabil, dan tersusun atas protein fibrosa. Sebagaian besar filamen intermediet berfungsi untuk menyokong sel dan inti sel. Letak filamen inibiasanya terpusat disekitar inti. Pada sel epitel, filamen intermediet membentuk anyaman yang berfungsi untuk menahan tekanan dari luar. Contoh filamen entermediet antara lain adalah kertin, vimentin, neurofilamen, lamina nuclear, dan keratin.
j. inti sel
Pada sel eukariotik, materi intinya telah diselubungi oleh suatu membran dan membentuk struktur inti sel atau nukleus.
Bagian –bagian yang menyusun inti sel antara lain adalah membran inti, pori membran,matriks inti sel (matriks), kromatin atau kromosom, dan anak inti (nukleolus).
Pada umumnya, inti sel berbentuk bulat, tetapi ada juga yang bentuknya seperti gelendong. Sel eukariotik umumnya memiliki satu inti sel, tetapi ada juga beberapa jenis sel yang memiliki inti lebih dari satu.
Berikut ini uraian tentang bagian-bagian penyusun inti sel.
1) Membran inti
Membran inti terdiri atas dua lapis, yaitu membran luar (membran sitosolik) dan membran dalam (membran nukleo-plasmik). Di antara kedua membran tersebut terdapat ruangan antar membran (perinuklear space) selebar 10 - 15 nm. Membran luar inti bertautan dengan membran ER. Pada membran inti juga terdapat enzim-enzim seperti yang terdapat pada membran ER, misalnya sitokrom, transferase, dan glukosa-6-fosfatase. Permukaan luar membran inti juga berikatan dengan filamen intermediet yang menghubungkannya dengan membran plasma sehingga inti terpancang pada suatu tempat di dalam sel.
2) Pori Membran Inti
Pada membran inti terbentuk pori-pori sebagai akibat pertautan antara membran luar dan membran dalam inti. Diameter pori berkisar antara 40 - 100 nm. Jumlah pori membran inti bervariasi tergantung dari jenis sel dan kondisi fisiologi sel. Fungsi pori membrane inti ini, antara lain sebagai jalan keluar atau masuknya senyawa – senyawa dari inti dan menuju inti, misalnya tempat keluarnya ARN – duta dan protein ribosom.
Pori membran inti dikelilingi oleh bentukan semacam cincin (anulus) yang bersama-sama dengan pori membentuk kompleks pori. Bagian dalam cincin membentuk tonjolan-tonjolan ke arah lumen pori. Pada bagian tengah pori terdapat sumbat tengah (central plug).
3) Matriks Inti (nukleoplasma)
Komponen utama dari matriks inti adalah protein vang kebanyakan berupa enzim dan sebagian adalah protein structural inti.
Matriks inti diduga ikut berperan dalam proses – proses pada materi inti, misalnya transkripsi, replikasi DNA, dan proses –proses lainva di dalam inti.
4) Materi Genetik
Bagian utama dari sebuah inti sel adalah materi genetik. Semua aktivitas di dalam sel dikendalikan oleh materi genetik. Pada waktu interfase, materi genetik dinamakan kromatin. Benang benang kromatin ini akan mengalami pemampatan (kondensasi) pada saat sel akan membelah. Kromatin yang mengalami kondensasi ini dinamakan kromosom. Hasil analisis kimia menunjukkau, bahwa kromatin tersusun atas DNA, RNA, protein histon dan protein nonhiston.
5) Anak Inti (Nukleolus)
Nukleolus banyak ditemukan pada sel-sel yang aktivitas . sintesis proteinnya tinggi, misalnya pada neuron, oosit, dan kelenjar. Di dalam inti, nukleolus tampak sebagai suatu struktur yang merupakan tempat pembentukan dan penyimpanan prekusor ribosom dan pembentukan sub unit ribosom. Selain itu, struktur ini merupakan tempat terjadinya proses transkripsi gen ARN ribosom (ARN-r).
k. Sentriol
Sentriol merupakan organel sel berbentuk silindris dengan diameter lebih kurang 2 pm (mikrometer) dan panjang lebih kurang 4 ptm. Di dalam setiap sel mengandun sepasang sentriol yang letaknya saling tegak lurus dekat inti sel. sentriol berfungsi sebagai bahan pembentuk sillia dan flagella , persis dengan sentriol. Jadi, selain sebagai komponen penyusun sentrosom, sentriol berfungsi sebagai tubuh basalis.
l. Silia dan Flagela
Kedua organel ini berfungsi sebagai alat pergerakan sel yang letaknya berada pada permukaan luar membran sel. Baik silia maupun flagella memiliki struktur yang sama, yaitu memiliki sumbu yang dinamakan aksonem. Struktur aksonem sangat kompleks karetra tersusun atas mikrotubulus dan protein. Jumlah silia pada umumnya banyak, sedangkan jumlah flagela hanya satu atau dua. Silia berukuran lebih halus dan lebih pendek dari pada flagela. Berbeda dengan sentriol, silia dan flagella dibungkus oleh membran. Membran silia dan flagela merupakan perluasan dari membran sel.
Contoh sel-sel bersilia adalah lapisan epitel saluran telur (oviduct) pada wanita, epitel saluran sperma (epididimis) pada lakilaki, pada organisme eukariotik uniseluler misalnya Paramaecium caudatum. Sedangkan flagela dapat ditemukan pada spermatazoa dan beberapa organisme eukariotik uni seluler misalnya Euglena viridis dan lain-lain.
Sel Eukariotik memiliki membran nukleus dan sistem endomembran.
Berikut struktur sel eukariotik:
1. Sel tumbuhan terdiri dari: nukleus, RE kasar, RE halus, ribosom, kompleks golgi, lisosom, membran plasma, mikrotubulus, mikrofilamen, mitokondria, badan mikro, kloroplas, vakuola, dinding sel.
2. Sel hewan terdiri dari: (membran nukleus, nukleoplasma+DNA, nukleolus) nukleus, vesikula, lisosom, RE kasar, RE halus, kompleks golgi, vesikula, sentriol, mitokondria, membran plasma, mikrofilamen, mikrotubulus.
diantara dua jenis sel yang secara struktural berbeda, sel prokariotik dan sel eukariotik. Hanya bakteri dan arkhea; alga hijau biru yang memiliki sel prokariotik. Sedangkan protista, tumbuhan, jamur dan hewan semuanya mempunyai sel eukariotik.